bioslawek
Dołączył: 09 Sty 2015
Posty: 96
Przeczytał: 0 tematów
Płeć: Mężczyzna
|
 Wysłany: Pią 17:25, 20 Sie 2021 Temat postu: Spór o nieredukowalną złożoność kaskady krzepnięcia krwi Wysłany: Pią 17:25, 20 Sie 2021 Temat postu: Spór o nieredukowalną złożoność kaskady krzepnięcia krwi |
|
|

Jednym z przykładów nieredukowalnej złożoności jest proces, który większość z nas w chwili skaleczenia się uważa za coś oczywistego - chodzi o krzepnięcie krwi. Kiedy byłem małym chłopcem i zdarzyło mi się skaleczyć to byłem przekonany, że krew na mojej ranie krzepnie, ponieważ ma kontakt z powietrzem. Uważałem że w kontakcie z nim wysusza się i dlatego robi się strup.
Karol Darwin być może miał podobne wyjaśnienie, kiedy obserwował tworzący się skrzep, ponieważ nie miał najmniejszego pojęcia o zawiłościach biochemicznych, które towarzyszą powstawaniu skrzepu tamującego upływ krwi. Być może gdyby miał choć część tej wiedzy na swoim naukowym koncie, to jego teoria nigdy nie ujrzałaby światła dziennego!
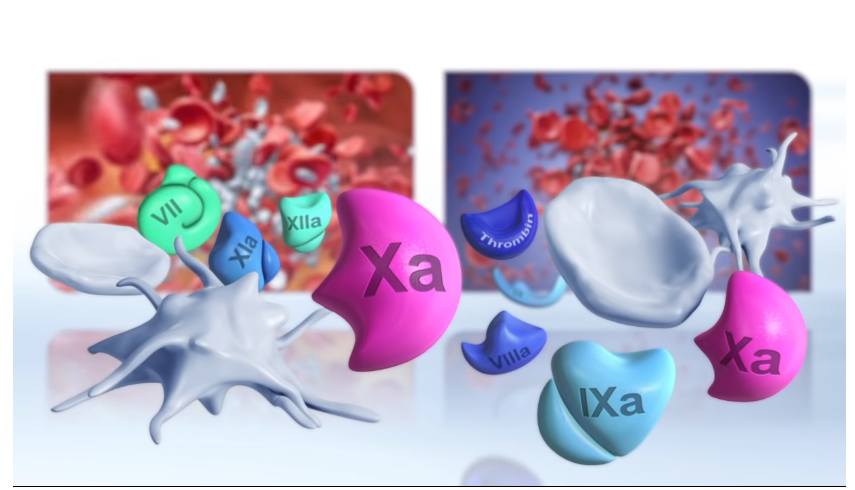
Normalnie dzieje się tak, że z przedziurawionego naczynia natychmiast zaczyna wypływać zawarty w nim płyn i cieknie dopóty, dopóki się ono nie opróżni. Jeżeli jednak przekłujemy czy przetniemy sobie skórę, upływ krwi szybko ustaje, gdyż tworzy się skrzep. Ale jak o tym dobrze wiedzą lekarze, krzepnięcie krwi jest bardzo złożonym, wielostopniowym procesem, w którym biorą udział liczne, oddziałujące na siebie białka. Uczestniczą one w tak zwanej kaskadowej aktywacji krzepnięcia. Ten delikatny proces leczniczy w ogromnej mierze zależy od tego, kiedy i z jaką szybkością zachodzą różne reakcje. Gdyby coś się źle potoczyło, człowiek mógłby się wykrwawić na śmierć albo przeciwnie, cała jego krew mogłaby ulec skrzepnięciu. Moment rozpoczęcia reakcji i ich szybkość mają więc żywotne znaczenie.
Z badań biologicznych wynika, iż w krzepnięciu krwi bierze udział wiele czynników i żadnego z nich nie może brakować. Chociaż istnieją różnice w składzie białek tworzących kaskadę krzepnięcia u pewnych gatunków, to zawsze konieczna jest minimalna ilość tych cząsteczek, która tworzy nieredukowalnie złożony rdzeń:

Biochemik Michael Behe zadaje pytanie: „Jak to się dzieje, że raz rozpoczęty proces krzepnięcia zostaje przerwany, zanim jeszcze cała krew (…) zamieni się w ciało stałe?” Wyjaśnia następnie, „iż tworzenie się skrzepu, wstrzymywanie dalszego krzepnięcia, wzmacnianie skrzepu oraz usuwanie go” stanowią zintegrowany system biologiczny. Jeżeli któryś element źle zadziała, cały system nie spełni swej funkcji.
Ewolucjonista Russell Doolittle, będący profesorem biochemii na Uniwersytecie Kalifornijskim, pyta: „Jakimże sposobem mógł powstać w wyniku ewolucji tak skomplikowany i precyzyjny proces? (…) Paradoksalna sytuacja: skoro każde białko musi zostać uaktywnione przez inne, to jak ten system mógł się rozwinąć? Jakiż pożytek przynosiła dowolna jego część, dopóki nie działała całość?” Doolittle stara się wyjaśnić pochodzenie tego procesu, posługując się ewolucyjną argumentacją. Jednakże profesor Behe zwraca uwagę, że „aby odpowiednie geny znalazły się na odpowiednich miejscach, potrzebny byłby niezwykle szczęśliwy traf”. Wskazuje także, iż wyjaśnienie podane w nonszalancki sposób przez Doolittle’a skrywa olbrzymie trudności.
Tak więc jednym z podstawowych zarzutów wobec modelu ewolucyjnego jest fakt, że nie wyjaśnia on pochodzenia tego, co prostsze być nie może, czyli nieredukowalnego rdzenia. Profesor Michael Behe oznajmia: „Chciałbym podkreślić, że dobór naturalny, będący motorem darwinowskiej ewolucji, działa jedynie wtedy, gdy istnieje coś, co może zostać wyselekcjonowane - coś, co przynosi pożytek już teraz, a nie dopiero w przyszłości”. No tak; dobór naturalny nie ma możliwości proroka  Nawet uczciwi przeciwnicy teorii inteligentnego projektu w przyrodzie przyznają, że darwiniści nie mają pojęcia, w jaki sposób układy niekompletne, które rzekomo były ewolucyjnymi prekursorami dzisiejszych systemów biochemicznych, mogły zdobywać przewagę selekcyjną: Nawet uczciwi przeciwnicy teorii inteligentnego projektu w przyrodzie przyznają, że darwiniści nie mają pojęcia, w jaki sposób układy niekompletne, które rzekomo były ewolucyjnymi prekursorami dzisiejszych systemów biochemicznych, mogły zdobywać przewagę selekcyjną:
Michael Behe to jeden z najlepszych biologów teoretycznych [ewolucyjnych] w naszych czasach. To nie bakteriolog typu PZ Myersa, dziennikarz rodzaju Dawkinsa czy genetyk populacji w typie Jerry Coyne’a. To wytrawny znawca ewolucjonizmu molekularnego, specjalista od struktury białek i prawdziwy naukowiec. Np. tutaj jest jego rewelacyjna praca z dziedziny biologii ewolucyjnej opublikowana w porządnej literaturze naukowej;
[link widoczny dla zalogowanych]

Jego wnioski o nieredukowalnej złożoności systemów biologicznych nie są wynikiem chciejstwa, tylko gruntownej analizy naukowej przy zastosowaniu tego samego aparatu poznawczego (teoretycznego), którym posługują się wszyscy inni biolodzy teoretyczni. Behe’ego można spotkać na każdej znaczniejszej konferencji poświęconej biologii ewolucyjnej. Jeżeli ktoś nie rozumie dlaczego mówimy o nieredukowanych sieciach genetycznych, to proszę poczytać, co pod inną nazwą niż nieredukowalna złożoność twierdzą sami darwiniści.
Autorzy artykułu w wikipedii bardzo się starali utopić istotę konsekwencji dla neodarwinizmu – fakt powszechnego występowania nieredukowalnej złożoności w biologii molekularnej i w efekcie wyszedł im niezły bełkot. To mieszanie genetyki populacyjnej z ewolucjonizmem molekularnym przypomina mieszanie wody z olejem. Dlatego też bardzo łatwo można odcedzić istotę problemu ukrywaną w słowotoku, zaprawionym różnymi sztuczkami retorycznymi, jaką jest fakt istnienia nieredukowalnej złożoności systemów biochemicznych. Jak zwał, tak zwał: nieredukowalna złożoność, czy silna negatywna epistaza, zawsze negatywna dla tendencyjnych „wyjaśnień” od których w tekście się roi oraz dla możliwości stopniowej ewolucji złożonych z wielu nierozerwalnie zintegrowanych i dopasowanych części maszyn molekularnych ?
[link widoczny dla zalogowanych]

[....]




Naukowcy wyeliminowali jeden gen i sieć genetyczna nie spełniła celu funkcji:




https://youtu.be/_qmNCJxpsr0
https://youtu.be/Ce2TjGY-r1o

[link widoczny dla zalogowanych]
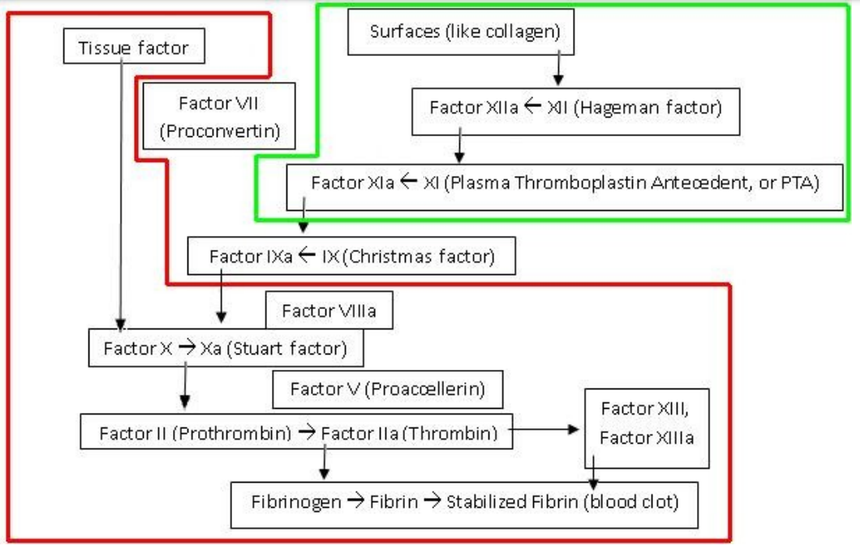
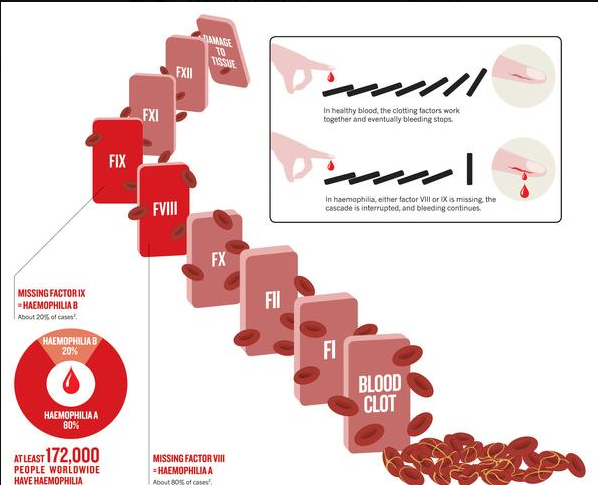
| Cytat: | | „Minoogowi, rybie bezszczękowej brakuje czynników IX i V . Są to ważne elementy kaskady krzepnięcia krwi, bez których mogą się obejść. Kiedy okazało się, że wielorybom i rybom kostnoszkieletowym brakuje wewnątrzpochodnego układu krzepnięcia krwi, Michael Behe stanowczo stwierdził: „ Tym gorzej dla nich ”. U ludzi brak czynnika IX powoduje zaburzenie krzepnięcia zwane hemofilią B , brak czynnika V powoduje również hemofilię, a brak obu z nich skutkuje ciężkom skazą krwotoczną. |

| Cytat: | Ryby kostnoszkieletowe i bezszczękowe mają zredukowany zestaw czynników krzepnięcia krwi w porównaniu do ssaków. Rybom bezszczękowym brakuje czynników IX i V. Czynnik tkankowy i czynnik VII bezpośrednio aktywują trombinę. Kaskada krzepnięcia jest inicjowana przez współdziałanie czynników IX i V.
Konsekwencje tego faktu dla „nieredukowalnej złożoności” są daleko idące.” |
[link widoczny dla zalogowanych]
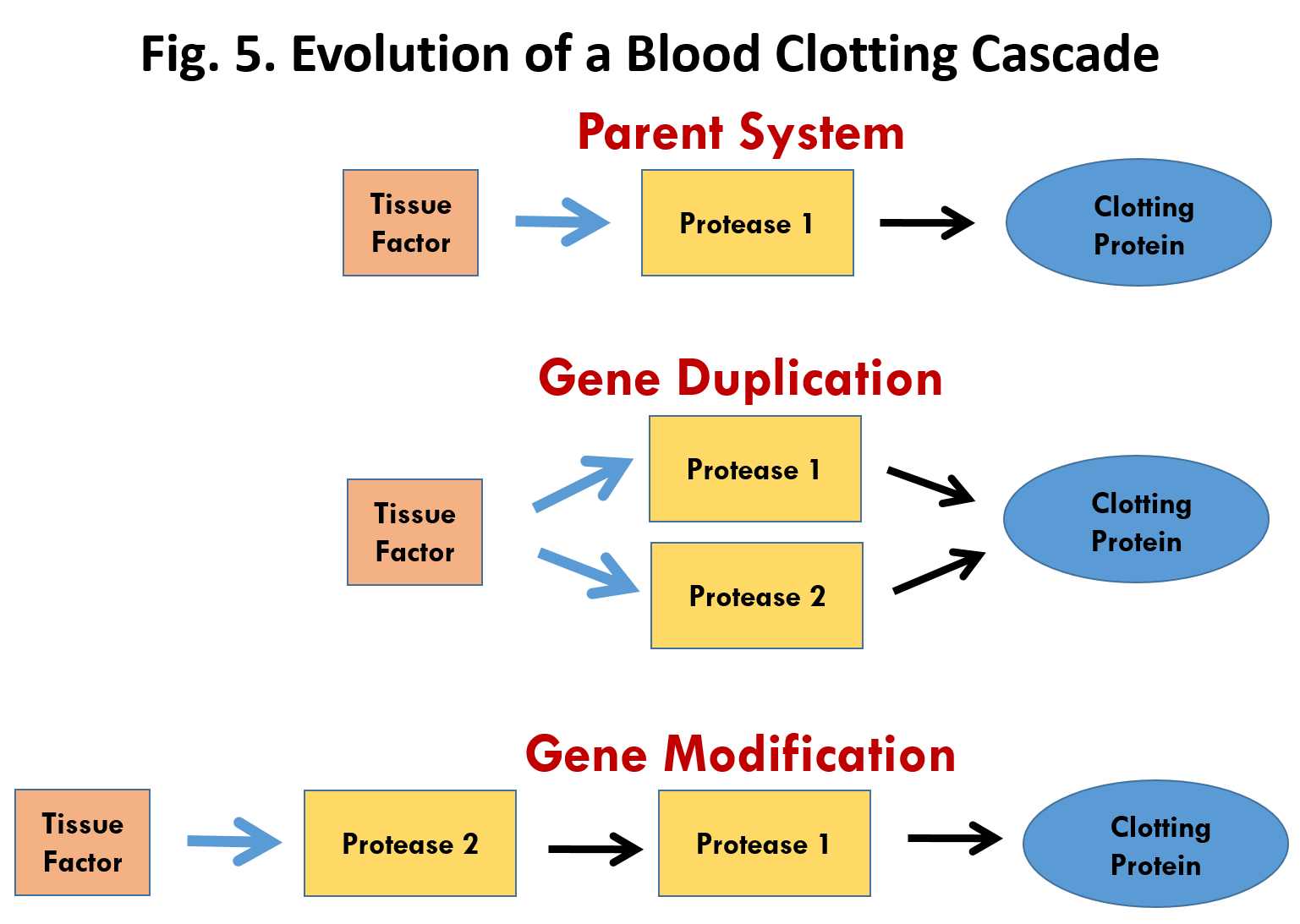



Russell Doolittle dużo pisze na temat postulowanych mechanizmów powstawania nowych genów [i zwiększających objętość genomu]. Takimi postulowanymi mechanizmami są duplikacje genów i tasowanie egzonów.
[link widoczny dla zalogowanych]
[link widoczny dla zalogowanych](Philosophical_Aspects_of_Origin)/Filozoficzne_Aspekty_Genezy_(Philosophical_Aspects_of_Origin)-r2005_2006-t2_3/Filozoficzne_Aspekty_Genezy_(Philosophical_Aspects_of_Origin)-r2005_2006-t2_3-s67-96/Filozoficzne_Aspekty_Genezy_(Philosophical_Aspects_of_Origin)-r2005_2006-t2_3-s67-96.pdf


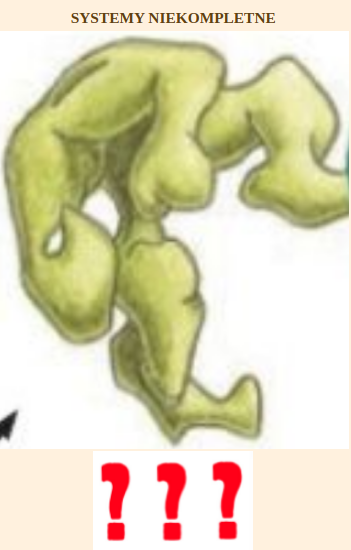
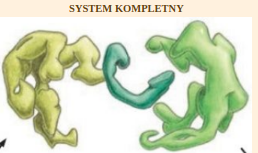
| Cytat: | | […..] Rozważmy [inny] hipotetyczny układ, w którym białka homologiczne do wszystkich części nieredukowalnie złożonego mechanizmu molekularnego z początku pełniły inne indywidualne funkcje w komórce. Czy nieredukowalny system mógł w takim przypadku zostać złożony z pojedynczych składników, które pierwotnie funkcjonowały osobno – jak proponują niektórzy darwiniści? Niestety, jak pisałem w Darwin’s Black Box, zarysowany powyżej obraz znacznie upraszcza ten problem […..] części układu molekularnego muszą automatycznie odnaleźć siebie nawzajem w komórce. Nie może ich ułożyć pewien inteligentny czynnik [……] Aby odnaleźć się wzajemnie w komórce, oddziałujące ze sobą części muszą mieć powierzchnie ukształtowane tak, żeby bardzo dobrze do siebie pasować [….] Pierwotnie jednak funkcjonujące z osobna składniki nie miałyby komplementarnych powierzchni. |

| Cytat: | Wszystkie oddziałujące ze sobą powierzchnie wszystkich składników musiałyby więc zostać dopasowane do siebie, zanim zaczęłyby działać razem. Dopiero wtedy mogłaby pojawić się nowa funkcja złożonego systemu. Dlatego mocno podkreślam, że problem nieredukowalności nie znika, nawet jeśli pojedyncze białka homologiczne do składników układu oddzielnie i pierwotnie pełniły swoje własne funkcje.
W kaskadzie krzepnięcia krwi jeden składnik wpływa więc na inny, który oddziałuje na następny i tak dalej. Argumentowałem, że kaskada jest nieredukowalnie złożona, ponieważ gdy usunie się jakiś jej składnik, ów proces albo natychmiast się włącza, albo definitywnie wyłącza. Na nic się zdaje – pisałem – postulat, że proces ten rozpoczął się od jednego czynnika, fibrynogenu, po czym dodano inne składniki, gdyż sam fibrynogen do niczego się nie przydaje. Nie warto też zaczynać nawet od czegoś w rodzaju fibrynogenu i niespecyficznego enzymu, który mógłby go rozszczepić, ponieważ krzepnięcie nie byłoby regulowane i możliwe, że czyniłoby więcej szkody niż pożytku. |

| Cytat: | Tak twierdzę ja. Jednak Russell Doolittle – wybitny biochemik zajmujący się białkami, profesor biochemii w University of California w San Diego, członek National Academy of Sciences, badający przez całe życie system krzepnięcia krwi – nie zgadza się ze mną. Doolittle napisał esej na sympozjum, dotyczący mojej książki i książki Richarda Dawkinsa Wspinaczka na szczyt nieprawdopodobieństwa.
Materiały sympozjum zostały opublikowane wydawanym przez Massachusetts Institute of Technology Boston Review. W eseju tym omawiane jest zjawisko duplikacji genu, dzięki któremu komórka może zaopatrzyć się w dodatkową kopię funkcjonującego genu. Doolittle wysunął przypuszczenie, że składniki procesu krzepnięcia krwi, z których wiele ma strukturę podobną do siebie nawzajem, powstały przez duplikację genu i stopniową dywergencję. Jest to rozpowszechniony pogląd wśród darwinistów (.......) To odwołanie się do duplikacji genu powtórzyło wielu naukowców recenzujących moją książkę, ale odzwierciedla ono powszechne nieporozumienie. Geny o podobnych sekwencjach sugerują tylko wspólne pochodzenie – nie mówią o mechanizmie ewolucji. Jest to istotny punkt mojego argumentu: świadectwo wspólnego pochodzenia nie jest świadectwem doboru naturalnego.(...) Pozwólcie,
że na zakończenie tej sekcji uwyraźnię to, że dwaj bardzo kompetentni naukowcy, profesorowie Miller i Doolittle, obaj wysoce umotywowani w dyskredytowaniu twierdzeń ID oraz gotowi przejrzeć całą biomolekularną literaturę w poszukiwaniu eksperymentalnych kontrprzykładów, obmyślają przykłady, które – gdy spojrzeć na nie sceptycznie – w rzeczywistości raczej wzmacniają niż osłabiają przypadek nieredukowalnej złożoności.
Oczywiście, nie dowodzi to, że twierdzenia nieredukowalnej złożoności są prawdziwe, czy że hipoteza ID jest poprawna, lecz pokazuje – jak sądzę – że naukowcy naprawdę nie wiedzą, jak potraktować nieredukowalną złożoność. Uwidacznia również, że teoria ID jest znacznie mocniejsza, niż uważają jej krytycy. |
| Cytat: | Wynika stąd także potrzeba traktowania z hermeneutyczną podejrzliwością darwinowskich scenariuszy, takich jakie zaproponowali Miller i Doolittle. Niektórzy naukowcy tak mocno wierzą w darwinizm, że wydają krytyczne osądy pod wpływem afektów i nieświadomie przeoczają dość oczywiste problemy scenariuszy darwinowskich lub ufnie utrzymują coś, co jest obiektywnie nieprawdziwe( .…) napotkałem gwałtowny sprzeciw – przeważnie ze strony internetowych entuzjastów darwinizmu, którzy pytali, po co w takim razie te setki czy tysiące artykułów badawczych opisujących darwinowską ewolucję nieredukowalnie złożonych systemów biochemicznych. Tworzyli oni strony internetowe, aby owe artykuły udokumentować.
[……]
Nie należy też pochopnie wnioskować o redukowalności systemów posiadających dodatkowe, czyli redundantne składniki, mogą one bowiem posiadać nieredukowalnie złożony rdzeń. Na przykład, samochód z czterema świecami zapłonowymi może jeździć z trzema lub dwiema świecami, lecz z pewnością nie pojedzie bez żadnej. Pułapki na szczury mają często dwie sprężyny w celu zwiększenia ich siły. |
| Cytat: | Taka pułapka może działać po usunięciu jednej sprężyny, ale nie będzie działała, gdy usunie się dwie.”
|
[link widoczny dla zalogowanych]
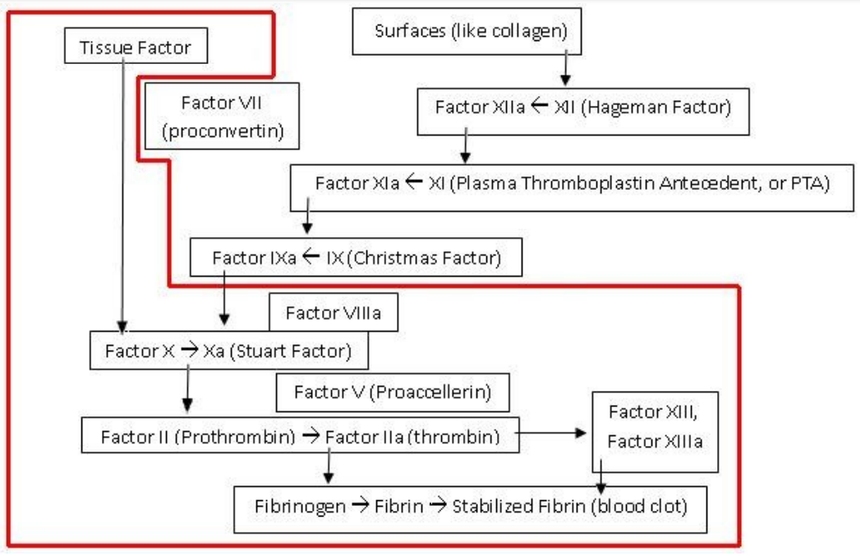
| Cytat: | „Kaskada krzepnięcia krwi
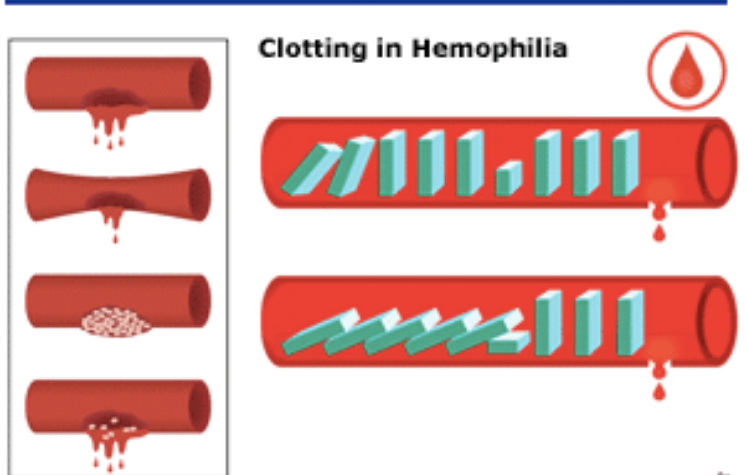
„Dr Miller wykazał też, że hipoteza o rzekomej nieredukowalnej złożoności kaskady krzepnięcia krwi została obalona eksperymentalnie już w 1969 roku, kiedy stwierdzono, że krew delfinów i wielorybów krzepnie mimo braku pewnej części kaskady, Wyniki te w roku 1998 zostały potwierdzone także na poziomie molekularnym. Niedawno opublikowany został zresztą raport z badań, zgodnie z którym u ryb najeżkokształtnych krew krzepnie pomimo braku nie tylko jednego, lecz trzech składników.(....)” |
[link widoczny dla zalogowanych]

Gdyby współczesnemu ssakowi lądowemu usunięto te trzy składniki, którym brakuje rybom bezżuchwowym (znokautowano geny, które je kodują), to czy mógłby przeżyć w przypadku krwotoku – czy dałoby się w ten sposób zredukować ssaczą kaskadę krzepliwości krwi do liczby mniejszej ilości elementów tak, aby dalej była funkcjonalna? Jak to się dzieje, że te ryby, czy inne wymienione przez Millera organizmy z prostszymi kaskadami krwi, przeżywają mimo braku pewnych składników biorących udział w kaskadzie krzepnięcia krwi u ssaków lądowych a te nie?
Od czego to jest zależnie: od biochemicznej organizacji tych różnych systemów - od specyficznego kontekstu molekularnego, czy od środowiska w jakim żyją te różne organizmy? Wniosek z tego taki, że innych rodzajów kaskad nie można traktować, jako FORM PRZEJŚCIOWYCH wiodących do ssaczego typu krzepnięcia krwi. Jedynie czego dowodzą, to wspólnego pochodzenia. Zostały opracowane na bazie podobnych projektów a nie wyewoluowały od wspólnego prekursora w postaci proto-kaskady krzepnięcia krwi.
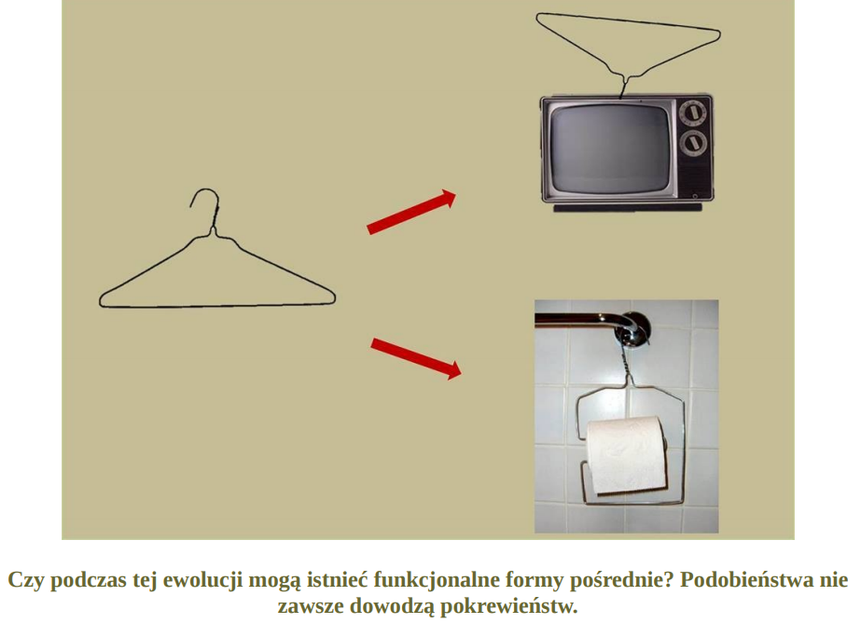

Wyobraź sobie, że do powstania jakieś niezbędnej korzystnej funkcji potrzebne są aż 4 mutacje: A,B,C,D. Że dopiero zestaw A,B,C,D może dać korzyść selekcyjną (dostosowanie). Jaka więc korzyść z pojedynczych mutacji: A, B , C czy D, skoro żadna z nich z osobna nie daje żadnej przewagi selekcyjnej? Każda z osobna jest neutralna, bezużyteczna? W tym przypadku, żeby cecha określana przez mutacje: A,B,C,D mogła dać przewagę selekcyjną, to te 4 [kompletny zestaw] mutacje musiałyby nastąpić za jednym zamachem, a prawdopodobieństwo takiego zdarzenia jest znikome. Jeżeli każda z tych 4 mutacji z osobna nie da przewagi selekcyjnej, to taka ewolucja po prostu nie ma szans nastąpić, ponieważ wtedy nie zadziała dobór kumulatywny.
https://youtu.be/wvTv8TqWC48
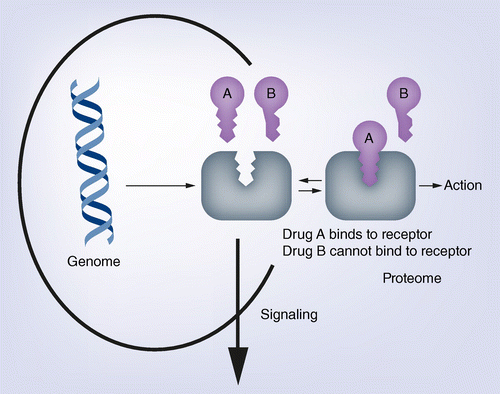
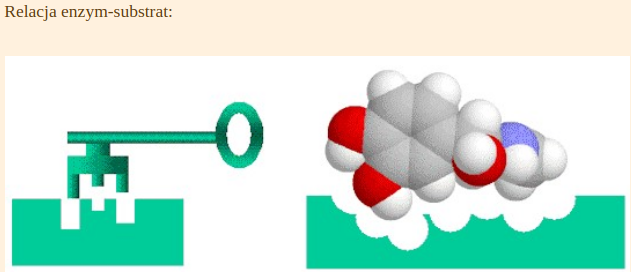
Istnieje wiele cech, których funkcję określają nierozerwalne zestawy konkretnie ulokowanych w genach nukleotydów. Jeżeli mutacja wprowadzi zmiany w takich konserwatywnych sekwencjach gen przestaje spełniać swoje funkcje i organizm ginie [efekt letalny]. Wiele enzymów, czy białek strukturalnych, posiadają takie konserwatywne domeny (centra aktywne) i jakiekolwiek zaburzenia paraliżują ich funkcję. W podobny sposób zachowuje się enzym zatruty jakimś antybiotykiem. Kontakt z toksyną powoduje, że jego centrum aktywne traci swoją funkcję i przestaje pasować, jak klucz do zamka do konkretnego substratu.
[link widoczny dla zalogowanych]
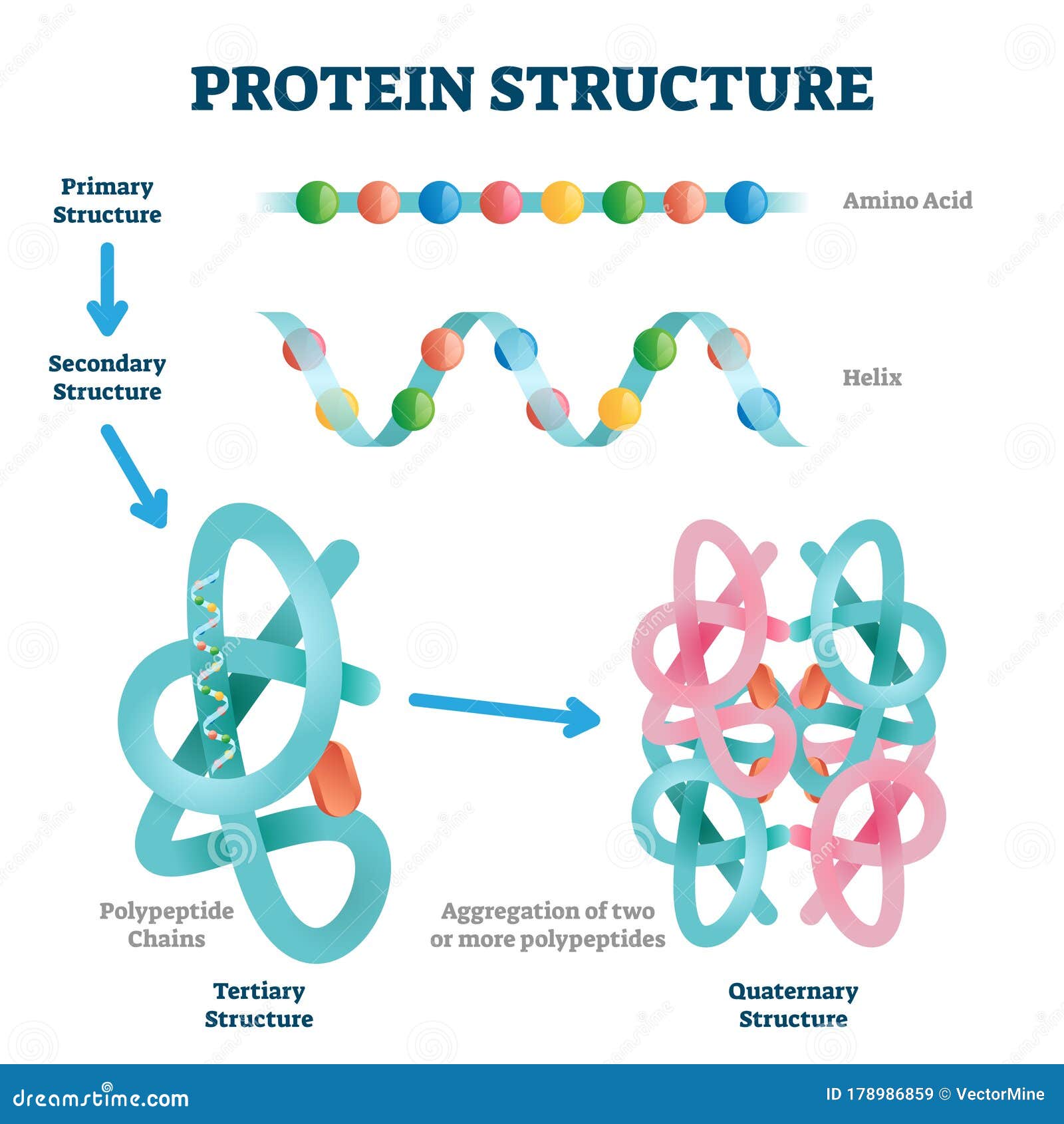
Polipeptyd, to długi łańcuch różnego rodzaju aminokwasów. Jedne z nich nie mają wpływu na formę (konformację, kształt) zwiniętego polipeptydu (białka) inne tak. Rozsiane i ulokowane w konkretnych miejscach wzdłuż łańcucha konkretne aminokwasy odpowiadają za:
-Odpowiednie zwijanie, które następuje według ściśle określonych sekwencji zdarzeń.
-Za powstawanie centrów aktywnych, kieszeni precyzyjnie dopasowanych do danego substratu.
-Za powstawanie komplementarnych domen pozwalających się łączyć z konkretnym typem białka.

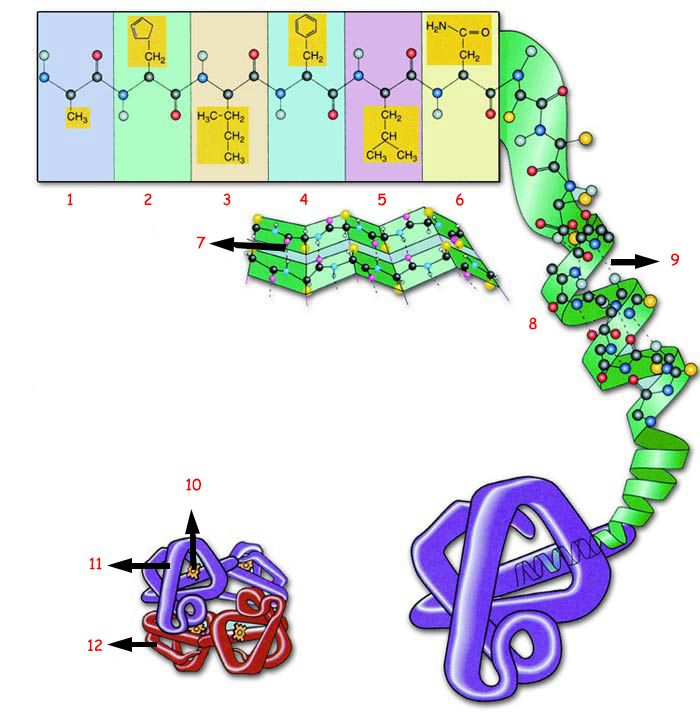
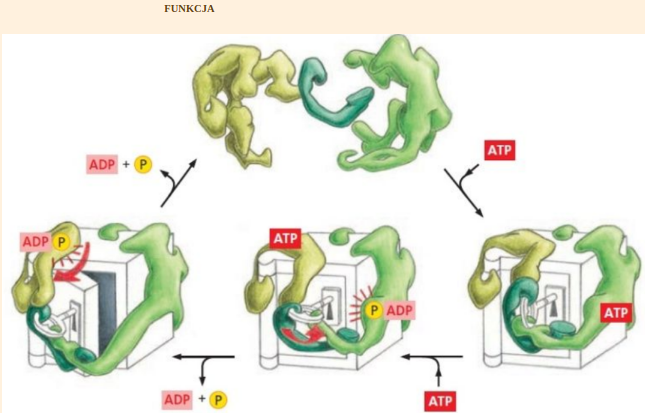
Jeżeli nie pojawiłby się komplet niezbędnych mutacji za jednym razem, tylko powstawałyby stopniowo na przestrzeni długich okresów czasu, to np. białko zwinęłoby się do pewnego momentu i ewolucja utyka w martwym punkcie. Tak też się dzieje w komórce. Zmutowane białka nie zwijają się prawidłowo i dlatego przestają być chronione przez inne białka, zwane chaperonami, ponieważ nie mają się jak przyczepić do źle sfałdowanego białka w celu chronienia go przed zniszczeniem. Wówczas specjalny białko zwane ubikwityną naznacza mutanta do kasacji. Następnie zostaje ono poddane recyklingowi, polegającemu na zmieleniu na poszczególne aminokwasy w specjalnej maszynie zwanej proteasomem:
Proteasom jest to wieloenzymatyczny kompleks utworzony z proteaz. Jest odpowiedzialny za degradację enzymów i białek regulatorowych. Zbudowany jest z cylindra 20S i dwóch regulatorowych kompleksów 19S znajdujących się na obydwu końcach cylindra. Podjednostka 19S rozpoznaje białka, które są przeznaczone do degradacji i odpowiada za jego rozwinięcie i skierowanie do cylindra. Natomiast cylinder jest odpowiedzialny za ich fragmentację. Proteasom degraduje tylko naznaczone wcześniej białka. Znacznikiem tym jest ubikwityna. Wchłonięte przez proteasom białko rozkładane jest do pojedynczych aminokwasów i krótkich peptydów złożonych z 10-12 aminokwasów.
https://youtu.be/JTR0XeRkyk0
https://youtu.be/hvNJ3yWZQbE
https://youtu.be/HIRz5hJf3mU
[link widoczny dla zalogowanych]

| Kod: | | „Cykl życiowy polipeptydu wewnątrz komórki. Po syntezie polipeptydu na rybosomie. Polipeptydy fałdują się w swoją unikalną trójwymiarową strukturę, obejmującą w niektórych przypadkach częściowo zwinięte związki pośrednie. |
| Cytat: | Te sekwencje pośrednie mają skłonność do agregacji z innymi białkami, jednak proces ten jest niekorzystny kinetycznie, co z kolei sprzyja tworzeniu się odkładających się i kumulujących zespołów toksycznych, które leżą u podstaw takich zaburzeń, jak choroba Alzheimera lub gąbczaste encefalopatie.
W celu wspierania prawidłowego fałdowania łańcucha polipeptydowego w jego pożądaną natywną strukturę, komórka jest wyposażona w zbiór asystentów fałdowania – chaperony, takie, jak białka szoku cieplnego (Hsp) lub białka opiekuńcze.
Białka niezdolne do osiągnięcia pożądanej konformacji trójwymiarowej, zużyte, lub nieprawidłowo sfałdowane są aktywnie degradowane przez komórkową maszynerię kontroli jakości, która obejmuje układ ubikwityna-proteasom (UPS).
Awarie w systemie proteasomów są powiązane z kilkoma zaburzeniami zwyrodnieniowymi. Wiadomo, że rybosomy pomagają we wczesnych stadiach fałdowania białek, co w praktyce oznacza, że bardzo niewiele sekwencji doświadcza procesu fałdowania ze stanu w pełni zdenaturowanego po syntezie w komórce – wiele polipeptydów fałduje się z częściowo zwiniętego.” |

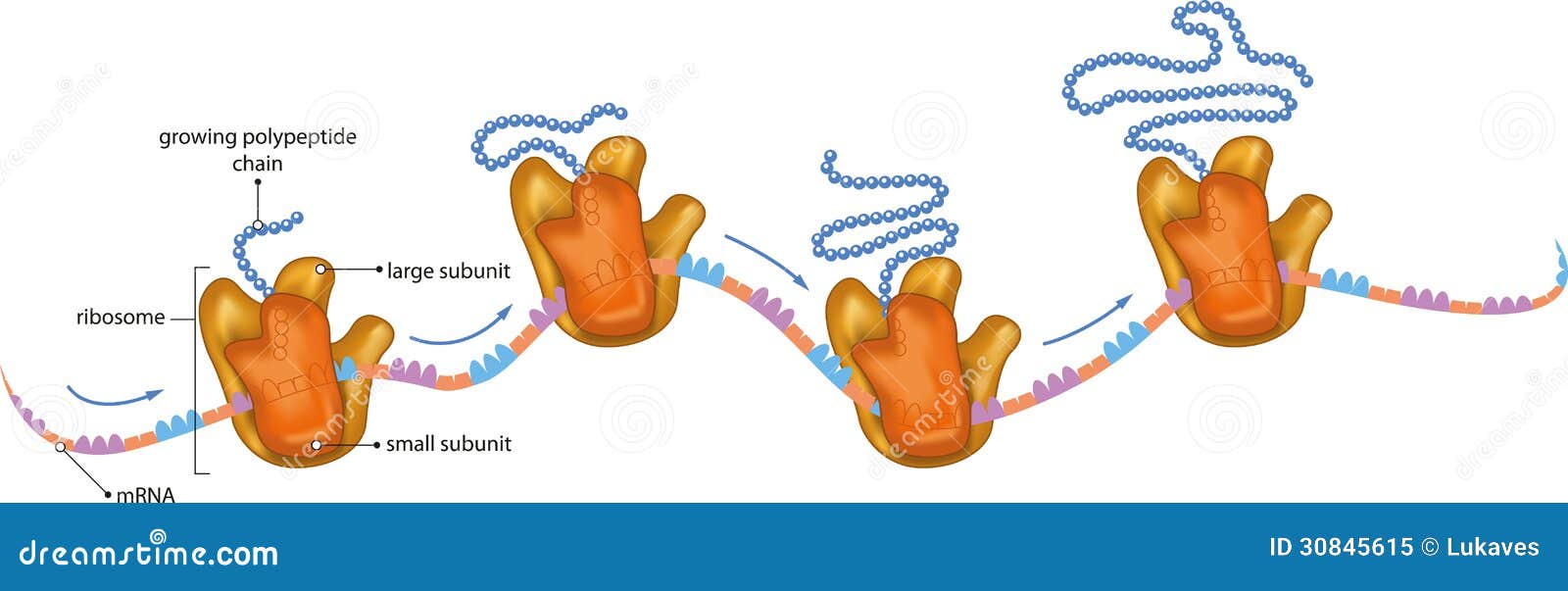
Niezbędne są też odpowiednie domeny komplementarnie, jak w przypadku puzzli, dopasowujące odpowiednie białka do siebie. Białka wchodzące w skład kaskady krzepnięcia to głównie proteolizy. Odpowiednie sekwencje pozwalające im ciąć jak piła inne białka byłyby bardzo niebezpieczne, gdyby nieodpowiednie dopasowanie domen pozwalało mieć kontakt sekwencji, którą można porównać do piły mechanicznej, z innymi białkami. Podobne sekwencje mają enzymy trawienne – każdy wie co się dzieje z białkami, kiedy dostaną się do żołądków. Podobnie wyglądałoby środowisko związane z układem krwionośnym, gdyby nie te subtelne dopasowania! Znając te fakty ogólnikowe twierdzenia, że kaskada wyewoluowała z układu trawiennego między bajki można włożyć.

Michael Behe opublikował piękny i madry artykuł, w którym dokładnie wyłożył w czym problem. Zatem darwiniści nie mają jedynie problemów z nieredukowalną złożonością na poziomie systemów biochemicznych, ale też na poziomie białek!
[link widoczny dla zalogowanych]





[link widoczny dla zalogowanych]



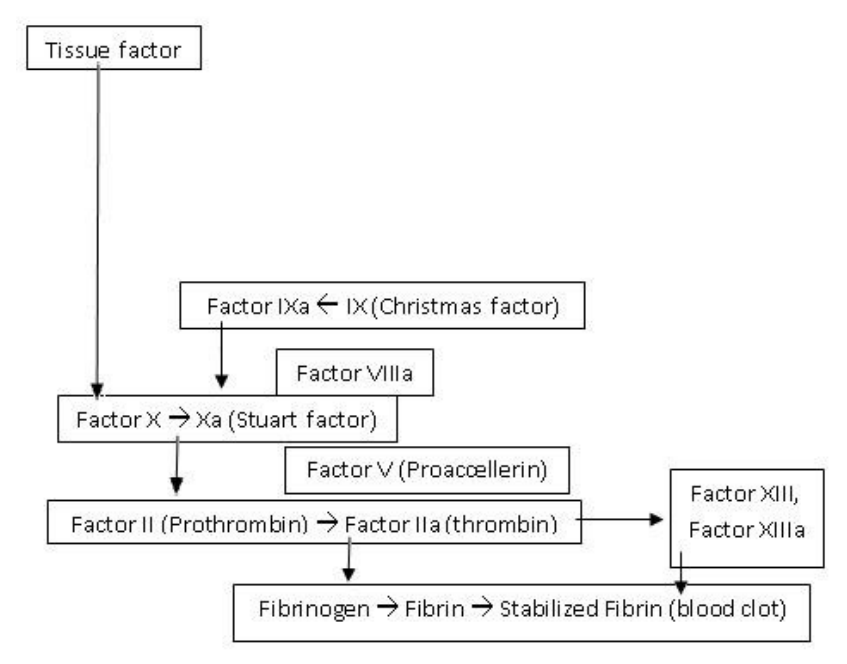

[link widoczny dla zalogowanych]

| Cytat: | | Schemat poglądowy głównych procesów tworzenia fibryny. Protrombina i trombina to białka aktywujące łańcuch reakcji chemicznych, które aktywują inne białka – fibrynogen. Monomery fibrynogenu tworzą nici fibrynowe. |

[link widoczny dla zalogowanych]

| Cytat: | | (A) Schematyczne przedstawienie protrombiny z jej składową domeną Gla, domeną kringle (K1 i K2) i domeną proteazy zawierającą łańcuch A i B. Miejsca aktywacji, R271 i R320, są zaznaczone na czerwono. Miejsca ataku proteolitycznego przez chymotrypsynę oznaczono na czarno. Rentgenowskie struktury krystaliczne zamkniętej (B, PDB ID: 6C2W) i otwartej (C, PDB ID: 5EDM) form protrombiny, które ulegają wzajemnej konwersji, wykorzystując elastyczność regionów łącznikowych, takich jak Lnk2. Forma otwarta jest atakowana przez chymotrypsynę w W468 w pętli autolizy. Zamknięta forma jest chroniona przed tym rozszczepieniem przez zapadnięcie się Y93 w kringle-1 do miejsca aktywnego, gdzie angażuje W468 i W547 w interakcjach układania. (D) Schematyczne przedstawienie czterech związków pośrednich konwersji protrombiny (ProT) do trombiny (T). Konwersja obejmuje rozszczepienie przy R271 i R320. Wcześniejsze rozszczepienie zrzuca pomocniczą domenę Gla i domeny kringle i generuje nieaktywną pośrednią pretrombinę-2 (Pre2). To ostatnie rozszczepienie oddziela łańcuchy A i B, które pozostają połączone wiązaniem dwusiarczkowym C293-C439, i generuje aktywną pośrednią meizotrombinę (MzT). |

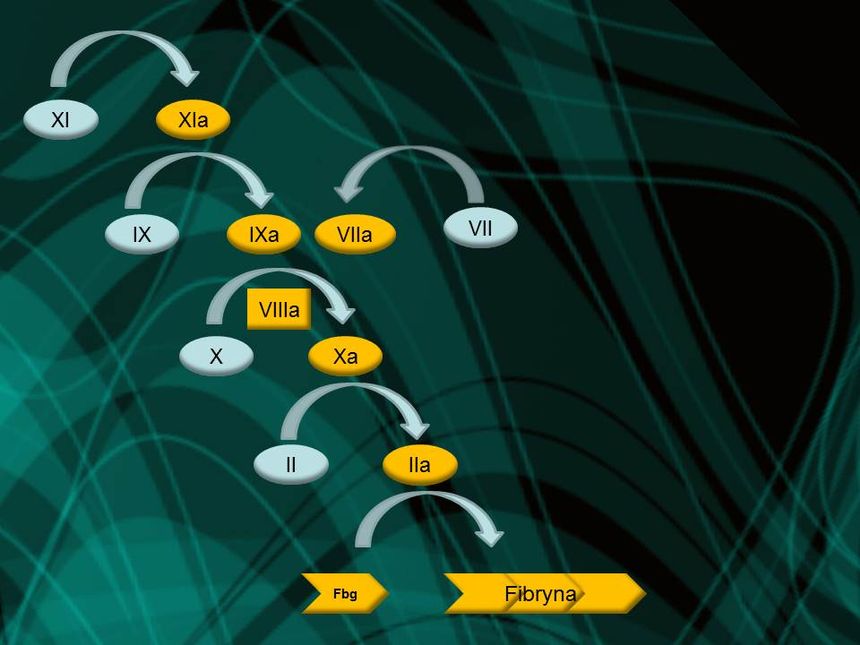
[link widoczny dla zalogowanych]


https://youtu.be/ly12vAUYcAU



[link widoczny dla zalogowanych]













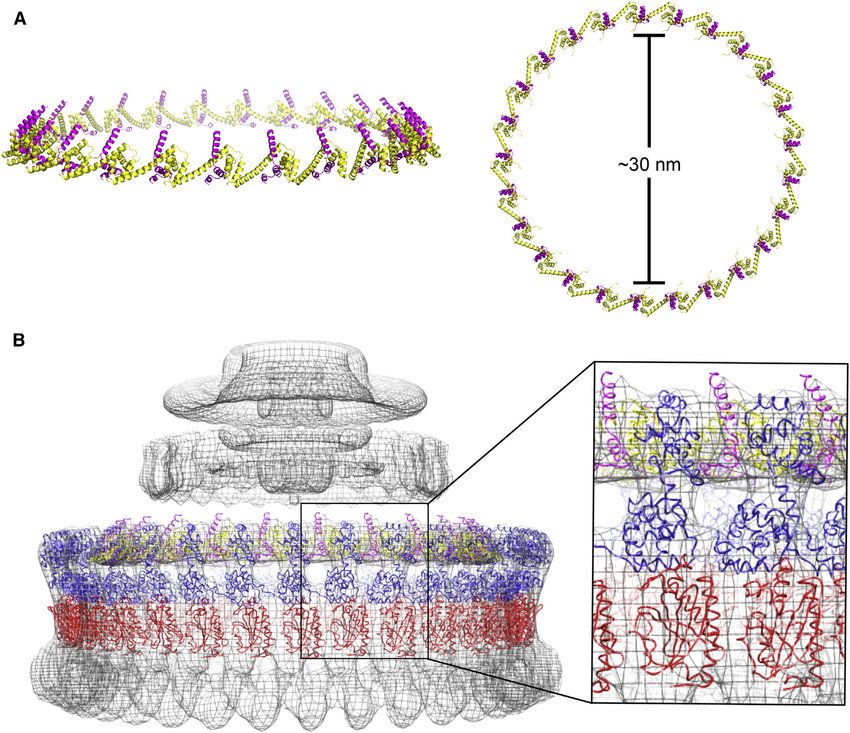
[link widoczny dla zalogowanych]



[link widoczny dla zalogowanych]


Ostatnio zmieniony przez bioslawek dnia Pią 17:32, 20 Sie 2021, w całości zmieniany 1 raz
|
|


